Heterochromatin is a crucial component of eukaryotic genomes, playing diverse roles in gene expression regulation, DNA replication, and repair. It is a highly condensed form of chromatin that is spatially segregated from euchromatin, typically located towards the nuclear periphery and surrounding the nucleolus. Despite its abundance in the nucleus, the mechanisms by which heterochromatin regulates and participates in genome organisation are still not fully understood. In this article, we will explore the functional role of heterochromatin in the formation of distinct chromatin compartments and how disruptions in heterochromatin formation can lead to genomic instability and disease.
The formation and maintenance of heterochromatin
Heterochromatin is categorised into two major types: constitutive heterochromatin (CH) and facultative heterochromatin (FH). CH refers to regions that are consistently silenced in all cell types and includes pericentromeric and telomeric repeated sequences, transposons, and some gene-poor regions of the genome. FH, on the other hand, consists of cell-type-specific heterochromatic regions that can switch into euchromatin under certain cues and is often found at developmental genes.
The formation, spread, and maintenance of heterochromatin involve complex molecular mechanisms. Histone modifications, such as methylation of histone H3 lysine 9 (H3K9me3), play a crucial role in heterochromatin formation. Histone Methyltransferases (HMTs), such as Suv39h in mammals, Su(var)3-9 in Drosophila, and Clr4 in yeast, catalyse the addition of methyl groups to H3K9, leading to the recruitment of heterochromatin protein 1 (HP1). HP1 recognises H3K9me3 and promotes the compaction of chromatin into heterochromatin domains.
The spreading of heterochromatin is facilitated by the dimerisation of HP1 and its interactions with various chromatin-modifying proteins and components of the nuclear envelope. This allows for the propagation of heterochromatin across large genomic domains. Additionally, non-coding RNAs, such as siRNAs, long non-coding RNA Xist, and major satellite repeat transcripts, play essential roles in recruiting the machinery necessary for heterochromatin formation.
Heterochromatin and genome topology
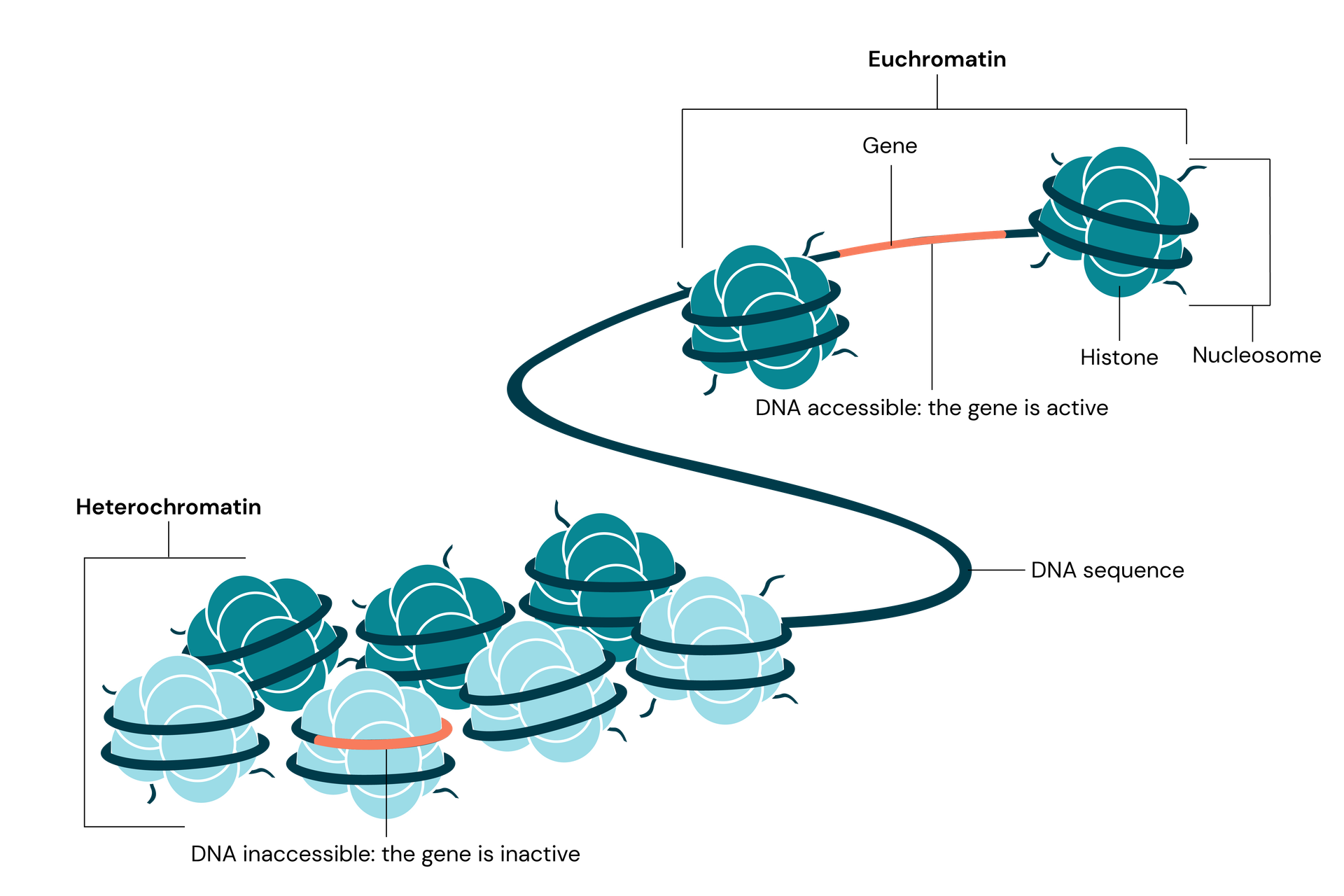
Advancements in Chromosome Conformation Capture technologies, particularly Hi-C, have provided valuable insights into the spatial organisation of the genome. Hi-C-based studies have confirmed that the genome is partitioned into chromatin compartments corresponding to euchromatin and heterochromatin. These studies have also led to the discovery of Topological Associated Domains (TADs), which play a crucial role in regulating gene expression by facilitating interactions between distant regulatory elements and genes.
The nuclear positioning of DNA sequences has been found to correlate with specific transcriptional states. Heterochromatin aggregates into discrete bodies within the nucleus and is often localised at the nuclear periphery. The repositioning of a gene from the nuclear periphery towards the interior of the nucleus can lead to changes in its expression levels.
Heterochromatin and gene silencing
The link between gene silencing and heterochromatin was first established through studies on X-ray-induced chromosome rearrangements in fruit flies. These rearrangements caused variegated phenotypes in the pigmentation of the flies’ eyes, resulting from the inactivation of the white gene without alterations in its sequence. It was later discovered that this inactivation was due to the relocation of the gene in proximity to a heterochromatic region, suggesting that heterochromatin could influence gene activity.
Transposable elements and silenced satellite repeated sequences have also been mapped to heterochromatic regions, providing further evidence of the repressive nature of heterochromatin. The formation of constitutive heterochromatin during cell differentiation and the distinction between constitutive and facultative heterochromatin have deepened our understanding of heterochromatin’s role in gene silencing.
Epigenetic regulation of heterochromatin formation
Epigenetic modifications, such as DNA methylation, can affect heterochromatin formation and gene expression regulation. DNA methylation is the addition of a methyl group to the cytosine residue of DNA, typically occurring at CpG dinucleotides. Methylation of CpG sites within heterochromatic regions can further stabilise the formation of heterochromatin and contribute to gene silencing.
The relationship between DNA methylation and heterochromatin formation is complex and regulated by various enzymes and proteins. The protein Methyltransferase 1 (Suv39h1) has been shown to bind to methylated DNA and stabilise its association with heterochromatin. Additionally, the binding of methylated DNA to HP1 has been found to promote heterochromatin maturation.
Publish
Disruption of heterochromatin and genomic instability
Dysfunction in heterochromatin formation and maintenance can lead to genomic instability and disease. Mutations in the genes responsible for heterochromatin formation have been implicated in various disorders, including cancer and developmental disorders. For example, mutations in the PRC2 complex, which deposits the H3K27me3 mark associated with facultative heterochromatin, have been linked to several types of cancer.
Furthermore, disruptions in the balance between heterochromatin and euchromatin can result in altered gene expression patterns and cellular dysfunction. Changes in the spatial organisation of the genome due to defects in heterochromatin formation can lead to misregulation of gene expression and impaired cellular processes.
Future directions and conclusion
While substantial progress has been made in understanding the role of heterochromatin in genome organisation, there are still many unanswered questions. Further research is needed to elucidate the molecular signals that trigger heterochromatin formation de novo and to identify new factors involved in heterochromatin remodelling. Additionally, the interplay between heterochromatin and other regulatory mechanisms, such as long-range interactions and epigenetic modifications, requires further investigation.
In conclusion, heterochromatin plays a critical role in genome organisation, gene expression regulation, and maintaining genomic stability. The formation and maintenance of heterochromatin involve complex molecular mechanisms, including histone modifications, non-coding RNAs, and interactions with other chromatin-modifying proteins. Disruptions in heterochromatin formation can lead to genomic instability and disease. Further research is needed to unravel the intricacies of heterochromatin function and its contribution to genome organisation in health and disease.